



随着厌氧消化理论研究的不断深入,厌氧消化工艺的研发和应用取得了迅速的发展,但处理效率低和!运行稳定性差是厌氧消化中普遍存在的问题,其中氨积累引发氨抑制是主要原因之一。文章简述了厌氧消化过程中氨抑制产生的机理及氨抑制的主要影响因素,介绍了氨抑制过程中微生物变化规律研究现状,总结了消除和缓解氨抑制的方法,并提出了厌氧消化氨抑制的重点研究方向。
厌氧消化作为一种可获得能源的可持续生物处理技术,在实际工程中得到了广泛应用。在厌氧消化过程中,氨抑制被认为是影响其整体效能的重要因素。虽然氮是厌氧消化系统中微生物新陈代谢所必须的元素,但是厌氧消化体系中过高的氨氮往往会抑制微生物的正常生命活动,尤其是产甲烷菌。
重点介绍了国内外厌氧消化氨抑制最新的机理研究,详细阐述了其主要的影响因素和消除措施,包括微生物驯化、pH 值调节、温度控制及 C/N 比调节等,为厌氧消化技术工程应用的未来研究提供一定的借鉴和参考。
厌氧消化作为一种能获得能源的可持续生物处理技术,其消化过程可以生成生物质能(主要为甲烷CH 4 ,含量为60% ~70%),是一种极具前景并可缓解气候变化的绿色能源 。
厌氧消化因具有稳定降解有机污染物并可在消化过程中产生可再生能源的特性,在对农业废弃物、食物残渣及污水厂剩余污泥等固体废弃物的无害化处理及资源化利用中得到了广泛应用 。在厌氧消化过程中,发酵物料内的含氮物质,如蛋白质、尿素和核酸等,在微生物的作用下最终被转化为氨氮(包括游离氨 NH3及NH+4) 。
尽管氨氮是厌氧消化过程厌氧微生物所必须的营养物,且一定浓度的氨氮可以为消化过程提供良好的缓冲作用,但是体系中过高浓度的氨氮对微生物有抑制作用,这被认为是导致厌氧消化反应系统崩溃的主要因素 。近年来,国内外对厌氧消化过程中氨抑制进行了大量研究,本文重点归纳了厌氧消化过程氨抑制的形成机制、影响因素以及应对措施,以期为厌氧消化反应系统的稳定运行提供一定的借鉴。
1 厌氧消化氨抑制机理研究
对厌氧消化过程中氨抑制形成机理的研究很多,但这些研究还不够完善,没有统一的共识。
厌氧消化过程通常包括物质溶解、水解、酸化、乙酸化及产甲烷五个步骤 ,故厌氧消化系统的稳定运行主要取决于水解发酵菌、产酸菌和产甲烷菌等微生物在正常生理活动下的协同作用,其中产甲烷菌对体系中氨浓度的耐受性最差 。
而氨抑制通常表现为稳定运行的厌氧消化体系中沼气产量下降以及挥发性脂肪酸 VFA 的积累 。许多研究者对氨抑制形成的机理提出了猜想,例如:产甲烷菌胞内 pH 变化、维持细胞正常生命活动所需的能量增加以及特定的酶促反应受到抑制等 。
在厌氧消化水溶液中,NH+4离子和游离氨(NH3)是氨存在的两种主要形式。游离氨由于其良好的渗透性,被认为是导致体系受到抑制的主要因素。
有学者通过纯菌种培养实验,推测体系中氨抑制对产甲烷菌的影响主要体现在以下两个方面:
1)NH+4离子可能直接抑制了甲烷合成所需酶的活性;2)疏水的游离氨分子可能通过被动扩散进入细菌细胞内,造成质子失衡或细胞内缺钾 。游离氨NH3经由被动扩散进入微生物细胞内,结合胞外质子 H+转化为 NH 4+,进而造成细胞内 pH 变化。
细胞为维持胞内质子平衡,通过细胞膜上钾泵消耗能量主动运输,将胞内钾离子移出至胞外,以维持胞内pH,由此增加了细胞维持能的需求并限制了一些特定的酶促反应。
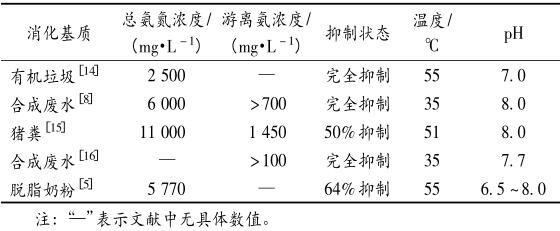
关于氨抑制阈值(见表 1)的研究较多,Hejnfelt和 Angelidaki 研究发现,在厌氧消化系统总氨氮浓度为 1 500 ~ 7 000 mg/L 内均有可能发生氨抑制现象。而不同的厌氧消化系统氨抑制阈值的差异主要受消化基质及接种物料特性、消化温度、系统内 pH及驯化时间等影响 。
表 1 不同厌氧消化系统氨抑制阈值
目前,对厌氧消化过程中氨抑制机理的研究,主要集中于体系中产甲烷菌的种群结构及多样性随氨浓度增加的变化。有学者研究表明,在厌氧消化过程中,氢营养型产甲烷菌较乙酸型产甲烷菌对体系中的氨浓度具有更强的耐受能力 。
Gao 等 通过设计餐厨垃圾厌氧消化反应器试验,结果发现,随着体系中氨浓度不断提升,试验最终阶段反应器内的辅酶F 420 (CoF 420 )的相对活性为最初阶段的 2. 4 倍,增加率达到 114%。
由于 CoF 420 的生化作用是低电位电子转移载体,特异性的将碳酸氢盐还原为氢气,进而被氢营养型产甲烷菌利用合成甲烷,从而证明随着反应器中氨浓度的提升,氢营养型产甲烷菌逐渐占据优势,这促进了特异性 CoF 420 的活性。
Niu 等利用16S rRNA 分子生物学技术研究氨氮对鸡粪高温厌氧消化过程中,产甲烷菌群落随体系中氨氮浓度不同的变化。
结果表明,氢营养型的甲烷热杆菌属(Methanothermobacter)由初始阶段的 9. 3% 发展至氨抑制阶段的 95%,大大超过乙酸营养型的甲烷八叠球菌属(Methanosarcina),在高氨氮体系的产甲烷过程中占主导地位。Demirel 及 Scherer 的研究也得出了相同的结论。
2 厌氧消化氨抑制影响因素及消除措施
氨抑制往往会给厌氧消化系统带来严重影响,甚至最终导致系统崩溃。因此,许多研究探讨了氨抑制的影响因素及其延缓或消除措施。主要包括微生物驯化、pH 值调节、温度控制、C/N 比调节等。
2. 1 微生物驯化
体系中高浓度氨氮可以直接抑制微生物活性,这往往是导致厌氧消化反应器失效的首要原因。研究表明,处理低氨氮浓度废水的厌氧消化反应器对系统中高氨氮浓度常具有更好的适应性。
通过逐步提高体系氨氮水平,对厌氧消化体系微生物进行驯化,可以提高产甲烷微生物种群对氨氮的耐受程度,是一种有效且经济的方式。
Abouelenien 等 通过对系统中微生物长达 254 天的驯化,最终实现体系在高氨氮浓度下产气量达到31 mL/g(VS)。Sung 和 Liu 的研究结果表明,尽管在高氨氮驯化下,体系中产甲烷菌整体的活性下降,但其对氨氮及 pH 变化的耐受性得到了强化。
Gao 等通过原位逐步进行氨耐受能力驯化,最终实现厌氧消化反应器在体系总氨氮达到4 275 mg/L 时依旧正常有效地运行。
尽管许多研究证实,通过驯化能够加强体系内微生物对不同氨氮浓度的适应能力,但其究竟是由于体系内已有微生物的代谢途径得到了转变,还是驯化出了能够适应高氨氮浓度的新生微生物群体,并未达成共识。
Gao 等 研究表明,在厌氧消化体系内总氨氮浓度从 2 341 mg/L(阶段1)变化至4 293 mg/L(阶段 5)的范围内,产甲烷杆菌(Methanobacteriaceae)相
对丰 度 由 阶 段 1 的 36. 16% 增 加 至 阶 段 5 的69. 73%,增长率达到 92. 84%。而产甲烷鬃菌科(Methanosaetaceae)及产甲烷球菌科(Methanosarcinaceae)相对丰度分别由阶段 1 的 31. 01% 和 26. 81% 降至8. 13% 和 2. 77%。在高氨氮浓度下,氢营养型产甲烷菌逐渐占据优势,而乙酸利用型产甲烷途径成为厌氧消化产甲烷过程的限速步骤 。
研究表明,当厌氧消化系统中氨浓度超过 3 g/L 时,体系中乙酸利用型产甲烷菌开始受到抑制,互养型乙酸氧化菌(SAOB)活性逐渐增强,其可将乙酸降解为氢气和二氧化碳,进而通过氢营养型产甲烷菌合成甲烷 。
Karakashev 等 研究也表明,随着体系中氨浓度升高,互养型乙酸氧化菌(SAOB)的活性逐渐超过乙酸利用型甲烷菌,在消耗乙酸的过程中占据优势。
2. 2 pH 值调节
厌氧消化体系的 pH 值不仅与微生物正常生命活动息息相关,且与体系中游离氨浓度具有紧密联系。由式(1) 可知,随着体系中 pH 上升,系统中NH+4会更多地转化为游离氨分子(NH3)。
而体系中产甲烷菌对氨浓度最为敏感,其活性首先受到影响,进而导致 VFA 积累,当其累积到一定程度时,体系中pH 随之下降,最终导致系统产气量下降,甚至崩溃。体系中 pH 值的有效控制对减缓氨抑制的影响至关重要 。

2. 3 温度控制
温度被认为是影响体系氨抑制阈值的显著性因素,其与厌氧消化系统中微生物生长速率及游离氨浓度(见式(1))联系密切。一般认为,随着温度升高,微生物新陈代谢速率加快的同时,也会导致体系中游离氨浓度相应增加。
此外,许多学者研究发现,对于高含氮有机废物,高温(50 ~ 60 ℃)厌氧消化较中温(30 ~40 ℃)厌氧消化更易受到氨抑制的影响,从而导致系统不稳定。
Hejnfelt 和 Angelidaki 对屠宰场废弃物进行厌氧消化,结果表明,高温厌氧反应器(55 ℃)最终体系总氮浓度达到 7 000 mg/L(游离氨浓度达到 999 mg/L)时受到抑制,而中温厌氧反应器(37 ℃)在游离氨浓度 达 到 400 mg/L 时 仍 稳 定 运 行。
Gallert 和Winter 通过设计两组生活垃圾厌氧消化反应器,反应温度分别为 37,55 ℃,当两组反应器分别达到50% 抑制时,其对应的游离氨浓度分别为 220,690 mg/L。然而,Masse 等 研究猪粪低温(10 ℃)厌氧消化,发现低温厌氧消化较高温及中温厌氧消化对系统高氨氮浓度耐受性更高,推测可能的原因是在低温条件下,体系游离氨浓度占总氨氮比值较低。
2. 4 C/N 值调节
对厌氧消化体系,如果发酵原料 C/N 比过高,会造成系统氮源不足,进而使碳源不能得到充分利用;反之,如果 C/N 比过低,则易导致系统氨积累进而对厌氧消化微生物产生抑制作用。选择不同特性的反应底物进行联合厌氧消化,用于减少体系受到氨抑制的风险,是一种经济有效且易于操作的措施。
调节发酵原料 C/N 比,可以有效控制系统中总氨氮浓度水平。但该过程较为缓慢,且只有在体系中产甲烷微生物群处于活性状态时,才能够起到相应的作用。如果在厌氧消化系统中的产甲烷微生物群已经受到严重抑制的情况下,此时调节发酵原料 C/N 比,系统也难以恢复。
De Vries 等 研究表明,相较于玉米青饲的单独厌氧消化,其与甜菜根及小麦酵母的联合厌氧消化,将生物能源丙三醇的产量提升了 568%。Hejnfelt和 Angelidaki 发现,在猪粪中加入 5% 猪肉副产品进行联合中温厌氧消化,其甲烷产量较猪粪单独厌氧消化提升了 40%。
Molinuevo-Salces 等通过往猪粪中投入一定的蔬菜加工废弃物进行厌氧消化,结果表明,体系中 VS 降解率得到了提升。
Shanmugam 和Horan 对制革废水进行厌氧消化,实验设计进料废水 C/N 比在 3. 2 ~30,结果表明,厌氧消化系统在系统 pH 值为 6. 5,C/N 比为 15 时,累积产气量达到最大,且此时系统游离氨浓度(817 mg/L)较低。相较于未调节 C/N 比的原废水,其游离氨释放减少了80%。
2. 5 其他影响因素及措施
许多学者研究发现,初始生物量、进料频率、加入膜接触器组件、微生物固定、微量元素添加及稀释等均对厌氧消化体系氨抑制有影响,通过采取一定的措施,可减缓氨抑制的影响。
Markou 等 研究螺旋藻厌氧消化,设计初始生物量分别为 125,250,500 mg/L,结果表明,厌氧消化系统初始生物量越低,越易受氨抑制影响。推测可能是由于生物量越高,其同化体系中氨氮的速率越快。
Tice 等 对微生物燃料电池进行研究,结果显示,在基底均为 2 g/L 乙酸钠的条件下,高进料频率(每 2天进料 1 次)组在系统总氨氮浓度达到 4 000 mg/L时,仍可以获得较高的功率密度(1. 1 ~ 1. 9 W/m 2 )。
而低进料频率(每 6 天进料 1 次)组在系统总氨氮浓度达到 2 500 mg/L 时所获得的功率密度更低。
通过提高进料频率,可以加强产电微生物对氨抑制的耐受能力。同时还指出,功率密度曲线相对于持续电流监控,能更有效地预测微生物燃料电池的氨抑制情况。
Lauterbock 等通过在屠宰场废弃物厌氧消化体系中加入中空纤维膜接触器组件用于持续去除系统中氨氮,结果显示,其游离氨浓度相较于对照反应器的1 000 ~ 1 200 mg/L,减少大约 70%。
系统更稳定的同时,取得了更高的甲烷产量及处理效率。在厌氧消化反应器中加入惰性负载材料(黏土、活性炭和沸石等),进行微生物固定,可以有效地减缓氨抑制,使消化过程更加稳定 。
Sasaki 等将碳素纤维纺织品(CFT)加入高温厌氧消化反应器进行产甲烷菌固定,结果表明,加入 CFT 的反应器在系统总氨氮达到3 000 mg/L 时,依然运行稳定。且通过 16S rRNA 基因分析,CFT 上附着大量的产甲烷菌。而未加入 CFT的对照反应器在系统总氨氮达到 1 500 mg/L 时,甲烷产率更低。
Banks 等 通过向厌氧消化系统加入微量元素硒和钴,并最优化其投加量,发现在最优条件下,可控制体系总氨氮浓度为 5 000 mg/L 并稳定运行,而对照组总氨氮浓度则达到 6 100 mg/L。
Kayhanian指出,稀释可以直接减轻厌氧消化系统的氨负荷,进而缓和氨抑制的影响。但由于稀释后会造成反应器体积浪费,处理效率降低,因此该方法并不经济 。
此外,通过一些物理化学方法,如化学沉淀 、吹脱 等,均可以达到去除厌氧消化反应器内氨氮,减缓氨抑制的目的。研究发现,两相厌氧消化系统较传统的单相厌氧消化系统,对系统氨氮具有更好的耐受性,同时可以提高系统的处理效率。
因此,可以考虑利用两相厌氧消化系统处理高含氮废物,以减轻消化过程中过高的氨氮对反应效率的影响。
3 结束语
目前,国内外针对厌氧消化氨抑制开展了大量的研究,但现有的研究更多的集中于氨抑制对产甲烷菌微生物群落的变化规律,且对于缓解氨抑制的措施还停留在实验室层面。
虽然发生氨抑制的厌氧消化体系仍可通过采取措施恢复,但该过程需要一定的时间进行复原,严重影响消化过程的连续性及效率,对于实际工程来说更是避之不及的。
因此,未来的研究方向应更多地集中在氨抑制过程中产甲烷菌与其他厌氧微生物的交互作用以及对体系氨抑制进行的预测、模拟等,在氨抑制早期对体系采取相应措施以避免氨抑制对系统带来的不良影响。




